*仅供医学专业人士阅读参考

胶质母细胞瘤(GBM)是一种侵袭性极强的癌症。
过往研究表明,表皮生长因子受体(EGFR)扩增是GBM中一个重要的致癌驱动因素[1-2]。然而,多项靶向EGFR的临床试验均未能取得良好疗效,表明EGFR扩增在GBM中的功能作用或有不同[3-4]。
据统计,约57.4%的原发性GBM伴随野生型EGFR(EGFRwt)基因的扩增,并且这与共同扩增的GBM特异性突变EGFRvIII(2-7号外显子缺失)相关[5-6]。有研究表明,EGFRvIII能协同触发配体诱导的或构成型激活的EGFR信号传导(在没有EGFR配体的情况下,由GBM中EGFR的表达导致自发二聚体化导致的信号传导[7])。
还有研究发现,构成型和配体诱导的EGFR激活触发了不同的信号通路[8-9]。目前,EGFR配体对于GBM发生发展的生物学功能,以及在GBM治疗中的临床实践意义是不清楚的。因此,构成型和配体诱导的EGFR激活在GBM中的作用和功能值得探索。
近日,来自德克萨斯大学西南医学中心的Amyn A. Habib教授团队,在《自然·细胞生物学》期刊上发表研究成果[10]。
他们发现,构成型激活EGFR通过激活TAB1-TAK1-NF-κB-EMP1通路促进GBM细胞侵袭。然而,配体诱导的EGFR激活通过上调桥接整合子3(BIN3),进而抑制DOCK7-RhoGTPase信号通路,导致GBM细胞增殖和侵袭能力下降。
简单来说,在EGFR扩增的GBM中,EGFR配体激活EGFR并抑制肿瘤细胞侵袭,将EGFR的作用从致癌基因转变为抑癌基因!郭高博士为论文的第一作者。

论文首页截图
让我们一起看看这项研究具体是如何开展的。
为了探索配体诱导的和构成型激活的EGFR及其介导的信号通路在GBM中的生物学作用。研究者在妙佑医疗国际患者来源的异种移植细胞系(PDX)和原代GBM神经球培养细胞中,检测了表皮生长因子(EGF)对GBM细胞侵袭的影响。
结果表明,添加EGF(EGFR配体)抑制了表达EGFRwt或EGFRvIII+EGFRwt的多个GBM细胞系的侵袭。使用siRNA敲降EGFR后,抵消了EGF对侵袭的抑制作用,表明GBM的侵袭作用是通过EGFR特异性介导的。

b:添加EGF,抑制GBM细胞侵袭;c:敲降EGFR后,抵消了EGF对侵袭的抑制作用
研究者也发现,将EGFR表达水平低的GBM细胞暴露于EGF会促进侵袭。然而,当EGFR在这些细胞系中过表达时,添加EGF又会抑制侵袭。
同时,使用不同剂量的siRNA敲降EGFR,随着EGFR表达水平的降低,EGF从抑制到促进侵袭逐渐过渡,这表明EGFR的表达水平,决定了配体诱导的EGFR信号传导的结果是促进还是抑制侵袭。
同时,研究者将GBM细胞注射到小鼠的大脑中,并通过体内活体显微镜实验证实,EGF抑制了GBM细胞的侵袭,并且EGF在高水平或低水平表达EGFR的多个细胞系中诱导细胞增殖。
然而,当siRNA敲降Src胶原同系物衔接蛋白(Shc)时,EGF诱导的增殖被抑制。Shc敲降抑制了响应EGF的丝裂原活化蛋白激酶1(ERK)激活,表明配体诱导的EGFR激活通过经典的Shc-ERK途径诱导增殖。
小鼠模型中EGF抑制了GBM细胞的侵袭
接下来,研究者进一步探索了EGFR抑制GBM细胞侵袭的下游信号机制。
RNA微阵列分析、WB和qPCR结果表明,BIN3是在添加EGF时上调程度最高的基因。重要的是,siRNA敲降BIN3阻断了EGF抑制GBM细胞侵袭的能力,同时过表达BIN3抑制细胞侵袭。
接下来,研究者利用质谱鉴定了与BIN3相互作用的蛋白,并找到了胞质分裂7(DOCK7)分子。
免疫共沉淀实验表明,EGF诱导了DOCK7和BIN3之间的相互作用,siRNA敲降DOCK7导致细胞侵袭性和RhoGTPase活性降低。
在DOCK7敲降的细胞中,添加EGF并没有进一步抑制侵袭或降低RhoGTPase活性,表明DOCK7位于BIN3的下游。同时,siRNA敲降BIN3抵消了EGF引起的RhoGTPase活性下调。
免疫共沉淀实验,EGF诱导了DOCK7和BIN3之间的相互作用
研究者也发现EGF诱导了GBM细胞中DOCK7-Y1118的酪氨酸磷酸化。DOCK7-Y1118F突变导致突变体不能与BIN3结合。结果表明,DOCK7的酪氨酸磷酸化以及EGFR上调BIN3对于DOCK7-BIN3相互作用同样重要。
随后,在无胸腺小鼠的颅内注射TGFα(EGFR配体)稳定表达的GBM12细胞或GBM9神经球细胞,结果表明,TGFα增加提高了小鼠存活率以及抑制了肿瘤侵袭能力。同时,在表达TGFα的肿瘤中,细胞增殖能力增强,但肿瘤体积明显减小。外源性输注EGF也会导致同样的结果。

b:TGFα增加提高了小鼠存活率;c:TGFα增加抑制了肿瘤侵袭
以上结果表明,配体诱导的EGFR激活通过上调BIN3,进而抑制DOCK7-RhoGTPase信号通路,导致GBM细胞增殖,侵袭能力下降。
紧接着,研究者探索了构成型EGFR信号通路在GBM细胞中的作用及背后机制。
研究者发现,上皮膜蛋白1(EMP1)是一个通过构成型EGFR信号上调的基因,siRNA敲降EMP1抑制了GBM中EGFR表达引起的细胞侵袭。
质谱分析表明,TAB1与TAK1可能参与了EGFR的构成型信号传导。并且,TAB1能够激活NF-κB[11]。siRNA敲降TAB1导致构成型EGFR介导的侵袭、TAK1的激活和NF-κB亚基p65磷酸化的丢失。siRNA敲降TAB1或TAK1也阻断了EGFR诱导的EMP1的上调,表明TAB1-TAK1-NF-κB-EMP1信号轴驱动了构成型EGFR诱导的细胞侵袭。
此外,研究者发现,在构成型激活的EGFR中观察到低水平的EGFR酪氨酸磷酸化,这有利于激活NF-κB-EMP1途径,促进细胞侵袭。而在配体诱导的EGFR中检测到高水平的EGFR酪氨酸磷酸化,这有助于激活EGR1-BIN3信号通路,抑制侵袭。
接下来,研究者发现Jak1和Jak3抑制剂Tofacitinib,在多个GBM细胞系中高度上调BIN3的水平,以及抑制细胞侵袭。Tofacitinib也能上调肝素结合的EGF水平(HB-EGF:EGFR配体),并诱导EGFR的高度激活。
同时,Tofacitinib阻断了STAT1和STAT3的磷酸化,而STAT3敲降模拟了Tofacitinib的作用,引起HB-EGF分泌增加、EGFR活化以及侵袭抑制。此外,西妥昔单抗能够完全抑制Tofacitinib诱导的EGFR活化、BIN3上调和侵袭抑制。
Tofacitinib在多个GBM细胞系中高度上调BIN3的水平并抑制细胞侵袭
研究者又探索了Tofacitinib对于小鼠颅内GBM肿瘤的影响,肿瘤细胞为GBM12TGFα的小鼠(高水平的EGFR配体),从Tofacitinib治疗中没有额外受益。然而,肿瘤细胞为GBM12V的小鼠(低水平的EGFR配体)在Tofacitinib治疗后,肿瘤侵袭能力下降,小鼠生存率显著增加。
此外,Tofacitinib也增加了小鼠肿瘤的增殖能力。稳定敲降HB-EGF的GBM39细胞对Tofacitinib治疗更敏感。这些结果表明,Tofacitinib可能对EGFR配体水平较低的EGFR扩增的GBM肿瘤更有效。
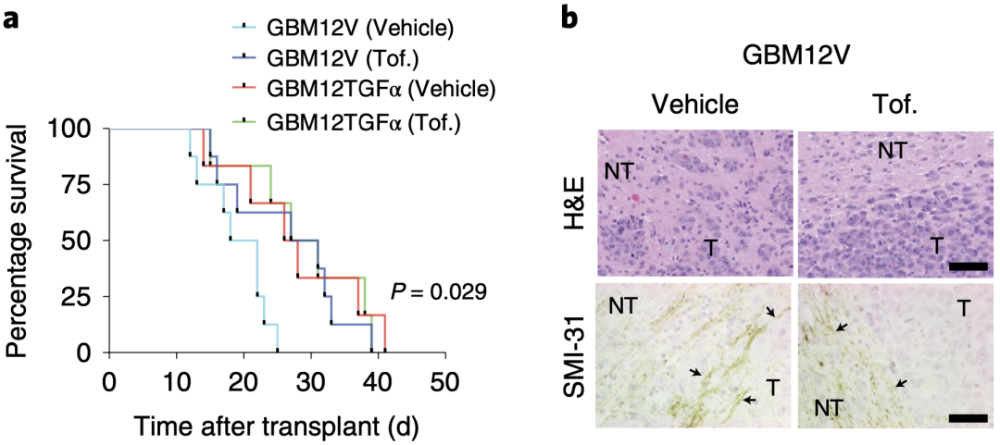
GBM12V和GBM12TGFα的小鼠在Tofacitinib治疗后生存情况,及肿瘤侵袭能力
在体内和体外实验中均证实了EGFR配体具有激活并逆转EGFR为抑癌基因的能力,为了在数据库中检测EGFR配体的表达情况,研究者进行了进一步的生信分析。
癌症基因组图谱(TCGA)数据显示,7个EGFR配体在GBM中有不同的表达,其中HB-EGF、TGFα和EGF是GBM中最丰富的配体,EGFR配体在非EGFR扩增的GBM中具有非常强的致癌作用(P=0.0009)。然而,在EGFR扩增的GBM中,EGFR配体具有显著的抑癌作用(P=0.0355)。
由于EGFR配体在GBM发生发展中表现出的双重作用,研究者进一步探索了EGFR配体及相关蛋白与GBM患者预后之间的关系。
结果表明,当GBM患者肿瘤内EGFR配体水平较低时,肿瘤细胞内EGFR的激活方式主要为构成型信号传导,而这部分患者的预后也较差。此外,相比于EGFR磷酸化水平较低的患者,EGFR磷酸化水平较高的患者,预后更好(P=0.0254)。并且,在36个GBM样本中,随着BIN3蛋白水平的升高,患者预后逐渐改善。
同时,为了了解EGFR及配体在肿瘤内是否存在区域表达的差异,研究者开展了酶联免疫吸附试验(ELISA)。
结果表明,在36个切除的GBM样本中,EGFR的表达在肿瘤区域之间没有显著差异,然而,HB-EGF在肿瘤中心部分的表达水平却更高,它也是这36个样本中最丰富的EGFR配体。更进一步的是,已有研究发现Sp1能促进HB-EGF的转录,Sp1的过表达上调了HB-EGF的表达水平[12]。同时,Sp1与HB-EGF共同定位于肿瘤组织相同的区域,这表明转录因子可能参与调节了EGFR配体的区域表达。
总的来说,这项研究表明在EGFR扩增的GBM中,EGFR配体激活EGFR并抑制肿瘤细胞侵袭,将EGFR的作用从致癌基因转变为抑癌基因,而这依赖于配体诱导的EGFR激活,上调BIN3并抑制DOCK7-RhoGTPase信号通路来实现。
这意味着,基于EGFR和EGFR配体水平的患者分层,可能有助于预测能从靶向EGFR中获益的患者亚群。

已购买课程的朋友,
直接进入小程序收听加餐哦~
参考文献
[1] Hatanpaa KJ, Burma S, Zhao D, Habib AA. Epidermal growth factor receptor in glioma: signal transduction, neuropathology, imaging, and radioresistance. Neoplasia. 2010;12(9):675-684. doi:10.1593/neo.10688
[3] Karpel-Massler G, Schmidt U, Unterberg A, Halatsch ME. Therapeutic inhibition of the epidermal growth factor receptor in high-grade gliomas: where do we stand?. Mol Cancer Res. 2009;7(7):1000-1012. doi:10.1158/1541-7786.MCR-08-0479
[4] Reardon DA, Wen PY, Mellinghoff IK. Targeted molecular therapies against epidermal growth factor receptor: past experiences and challenges. Neuro Oncol. 2014;16 Suppl 8(Suppl 8):viii7-viii13. doi:10.1093/neuonc/nou232
[5] Roos A, Dhruv HD, Peng S, et al. EGFRvIII-Stat5 Signaling Enhances Glioblastoma Cell Migration and Survival. Mol Cancer Res. 2018;16(7):1185-1195. doi:10.1158/1541-7786.MCR-18-0125
[6] Jahani-Asl A, Yin H, Soleimani VD, et al. Control of glioblastoma tumorigenesis by feed-forward cytokine signaling. Nat Neurosci. 2016;19(6):798-806. doi:10.1038/nn.4295
[7]Endres NF, Das R, Smith AW, et al. Conformational coupling across the plasma membrane in activation of the EGF receptor. Cell. 2013;152(3):543-556. doi:10.1016/j.cell.2012.12.032
[8] Chakraborty S, Li L, Puliyappadamba VT, et al. Constitutive and ligand-induced EGFR signalling triggers distinct and mutually exclusive downstream signalling networks. Nat Commun. 2014;5:5811. Published 2014 Dec 15. doi:10.1038/ncomms6811
[9] Guo G, Gong K, Wohlfeld B, Hatanpaa KJ, Zhao D, Habib AA. Ligand-Independent EGFR Signaling. Cancer Res. 2015;75(17):3436-3441. doi:10.1158/0008-5472.CAN-15-0989
[11] Hirata Y, Takahashi M, Morishita T, Noguchi T, Matsuzawa A. Post-Translational Modifications of the TAK1-TAB Complex. Int J Mol Sci. 2017;18(1):205. Published 2017 Jan 19. doi:10.3390/ijms18010205
[12] Miyata K, Yotsumoto F, Nam SO, et al. Contribution of transcription factor, SP1, to the promotion of HB-EGF expression in defense mechanism against the treatment of irinotecan in ovarian clear cell carcinoma. Cancer Med. 2014;3(5):1159-1169. doi:10.1002/cam4.301
责任编辑丨BioTalker